
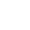

头条新闻
当前位置:首页 > 新闻公告 > 头条新闻
上海光源三项用户成果接连在《自然》发表
时间:2012-03-14
一)、新型铁基超导体研究中取得重要进展
2010年发现的新型铁基硫族化合物超导体A1 - xFe2 - ySe2(其中 A = K, Rb 或 Cs, 及部分替代的Tl)为非常规超导体的研究探索开辟了新的研究方向和提供了新的机遇。这类超导体在常压下的超导转变温度约为32K,中子散射研究结果表明,这类超导样品具有很强的反铁磁磁距并在铁离子的正方型格子中存在铁离子有规律的缺位。为何这样高TC的超导电性能够在如此强的磁背景中存在,以及反铁磁和铁离子有规律的空位如何对超导电性产生影响等问题倍受关注。大量的理论和实验研究结果表明,对于关联电子系统中的超导电性是由其晶体结构、电荷、轨道和自旋的状态及其相互作用所决定的,而这些因素对超导电性的影响可以通过外部参数的改变,如施加压力、磁场等进行调控。其中,压力是一种“干净”和有效的调控方法,它的独特之处在于不用改变研究系统的化学构成就能实现对系统的电子结构和晶体结构及其相关合作现象的有效调控,从而揭示其内在的物理机制。比如,利用压力手段可以改变电子密度、电子轨道的杂化和磁性能等,由此导致许多重要物理现象,如金属——绝缘体相变、价态变化、超导相的出现等。
最近,中国科学院物理研究所国家超导实验室赵忠贤院士小组的孙力玲研究员及其合作者与美国卡内基研究院地球物理实验室毛和光院士、陈晓嘉博士等合作,利用自行研制的高压——低温——磁场联合测试系统对这类新型铁基硫族化合物超导体进行了系统的高压原位输运性能和磁性能的研究。他们发现这类超导体的超导转变温度在压力小于10 GPa (1GPa=1万大气压)时随着压力的升高而逐渐降低,直至消失;而当压力高于10 GPa以后,系统出乎意料地进入了一个新的超导态,超导电性再次出现。这个由压力诱发的第二个超导相的超导转变温度高达48K,远远高于第一个超导相的最高超导转变温度,是已有报导的铁基硫族化合物超导体家族中的最高超导转变温度。为了澄清压力驱动的超导再现是否与压致晶体结构的变化有关,在与上海光源硬X射线微聚焦线站(BL15U1)杨科、李爱国的紧密合作下,在高压下对这种材料的晶体结构进行了X-射线微区衍射测量和分析,得到了晶体结构与压力间的直接实验证据。原位的X-射线微束衍射实验结果表明,在该研究的压力范围内没有发现一级结构相变,这表明压力诱发的超导再现是在同结构下产生的。该项研究中发现的同结构下由压力调控的超导电性的消失和随后的超导电性的再现现象在铜氧化物和铁基高温超导体中均未曾发现过,因此,这一结果不仅对理解这类新型铁基硫族化合物超导体的超导机制和探索新的超导体提供了启迪,同时,也为进一步开展高温超导体超导机理的研究提出了新的研究课题。该项研究结果发表在Nature 483, 67–69 (01 March 2012)。此前, Nature网站的新闻专栏对该项成果进行了专题报导。
http://www.nature.com/news/superconductor-breaks-high-temperature-record-1.10081
http://www.nature.com/news/superconductor-breaks-high-temperature-record-1.10081

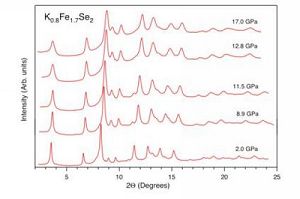
图1 压力下新型铁基硫族化合物超导体Tl0.6Rb0.4Fe1.67Se2、 K0.8Fe1.7Se2 和K0.8Fe1.78Se2的超导相图。图中展示了两个超导相的超导转变温度与压力的依赖关系。 所有的样品在压力下均具有两个超导相存在区。其中SC-I表示超导体第一个超导相存在区, SC-II表示超导体在压力诱发下再现的第二个超导相存在区,NSC表示非超导相。 第二个超导相的最高超导转变温度高于第一个超导相的最高转变温度。在压力为12.5 GPa时, TC=48 K, 这是目前新型铁基硫族化合物超导体的最高转变温度。
图2 不同压力下K0.8Fe1.7Se2 的X射线微束衍射图谱
二)、揭示植物感受紫外线B波段的分子机理
2012年3月1日,清华大学施一公教授研究组、北京大学邓兴旺教授研究组合作在Nature在线发表了题为“Structural basis of ultraviolet-B perception by UVR8”的论文,解析了植物拟南芥感受紫外线B波段(280 - 315 nm)的光受体UVR8的晶体结构,并对其感光机理做出了解释。
在植物的生长过程中,光发挥着极其重要的作用,包括提供能量,调控植物生长、发育的各个阶段(例如发芽、开花等)。植物对于光的感受,是通过一类叫做光受体的蛋白执行的,光受体感受光信号,再把信号传给下游调控因子,从而使植物做出相应反应。对光受体感光机理的研究,吸引了众多生物学家的目光。植物的红光/红外光受体、蓝光受体发现较早,目前都有比较深入的功能和结构方面的研究。但是紫外线B波段的光受体,在2011年4月才被鉴定,研究人员发现,在拟南芥中,一个名为UVR8的蛋白正是人们寻找了许久的紫外线B波段的光受体。生化实验为其感光机理提供了一些线索,表明紫外线(280 - 315 nm)照射会使UVR8从二聚体变为单体,但是具体分子机理的阐明,还有赖于高分辨率的结构。
为了解决这个问题,施一公与邓兴旺的研究组合作,利用上海光源生物大分子晶体学线站(BL17U1)解析了UVR8两个突变体的高分辨率晶体结构(1.8 Å - 2.0 Å),对相关分子机制的阐明起到了关键作用。UVR8在紫外线B波段光照之前,是一个由两个相同的单体形成的二聚体,其中每个单体由7个富含β-折叠的WD40结构域组成。两个单体通过众多氢键紧密结合在一起。通过野生型与突变体UVR8结构的比对,以及进一步的基于结构的生化分析,作者阐明了UVR8的感光机理:UVR8单体中,两个色氨酸(W285和W233)组成了感受紫外线B波段的核心基团,当受到紫外线(280 - 315 nm)照射时,两个色氨酸的吲哚环电子被激发,破坏了W285和W233与相邻的两个精氨酸R286、R338之间的紧密的电荷作用(cation-π interaction),影响了R286、R338在结构中的稳定性,而R286、R338正是参与形成两个UVR8单体间氢键的关键氨基酸,二者受到扰动,破坏了相应氢键,从而使UVR8二聚体解聚。
这些成果为研究人员在分子水平理解植物感光机理提供了帮助,也为进一步的计算机模拟、生物物理学研究奠定了基础。
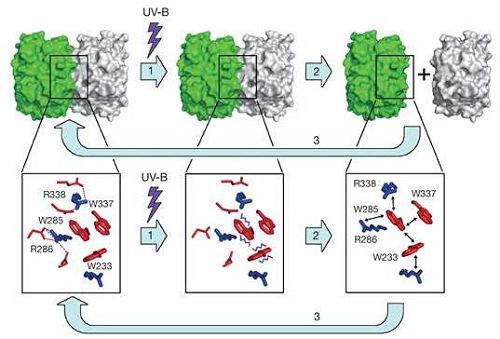
图为基于UVR8晶体结构推测的感受紫外线B波段的分子机理
三)、谷氨酸:γ-氨基丁酸(GABA)反向转运蛋白工作机理
2012年3月11日,清华大学生命学院施一公教授研究组在Nature在线发表了名为”Structure and mechanism of a glutamate-GABA antiporter”的科研论文,报道了大肠杆菌谷氨酸:γ-氨基丁酸(GABA)反向转运蛋白(GadC)的晶体结构,并结合生化实验提出了GadC转运底物的可能机制。
2011年在欧洲爆发的肠溶血性大肠杆菌疫情导致数千人感染,多人死亡,并引起了极大的社会恐慌。大肠杆菌O104:H4菌株是这一疫情的罪魁祸首。食物来源的大肠杆菌必须通过极酸的胃环境(pH~2)才能到达肠道,为了保证在如此低的pH下仍然能够存活,大肠杆菌进化出了多个抗酸系统来对抗极酸环境。因此研究大肠杆菌抗酸机制对人类健康有直接的重要意义。
大肠杆菌抗酸系统II(Acid Resistance system 2)通过谷氨酸: γ-氨基丁酸反向转运蛋白(GadC)将细胞外的谷氨酸转运到细胞内,在胞浆内谷氨酸发生脱羧反应,消耗一个质子生成γ-氨基丁酸,而产物γ-氨基丁酸再被GadC转运到细胞外。整个过程相当于向细胞外排出一个质子,降低了细胞内的质子浓度,从而起到了抗酸的作用。理解GadC工作机理对于研究肠道致病菌抗酸系统十分关键。
利用从上海光源生物大分子晶体学线站(BL17U1)收集的晶体衍射数据,施一公教授研究组解析了大肠杆菌GadC母体晶体的高分辨率的晶体结构。结构分析表明,含有12个跨膜螺旋的GadC在pH碱性条件下呈现出转运通道开口朝向胞内的构象,令人惊奇的是,GadC羧基端结构域在细胞内一侧像塞子一样将转运通道封闭住。同时进一步的生化实验表明,GadC对底物的转运严格依赖于环境pH值:野生型GadC在pH小于6.5的环境下才具有转运能力,而在pH大于6.5的环境中完全没有活性。羧基端“塞子”结构域在GadC感受pH的过程中起到了重要的调节作用。这样一种机制既保证了抗酸系统在极酸环境中能够启动并转运底物,又防止其在正常生理条件下造成胞内质子不必要的外流。
以上研究成果为进一步研究大肠杆菌抗酸机制提供了重要线索。这是施一公教授研究组继2009年、2010年报道Arginine:Agmatine反向转运蛋白AdiC的结构与机理之后,在肠道细菌抗酸性研究中的又一重要成果。
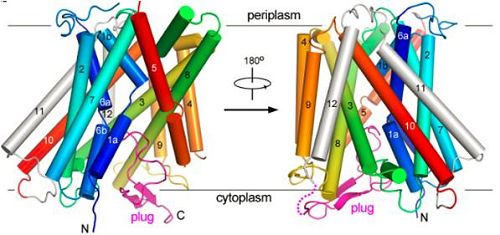
图为大肠杆菌谷氨酸: γ-氨基丁酸(GABA)反向转运蛋白 (GadC) 的晶体结构